Plants, like most animals, have a hierarchical organization consisting of organs, tissues, and cells. The basic morphology of vascular plants reflects their evolutionary history as terrestrial organisms that inhabit and draw resources from two different environments – below the ground and above the ground. The ability to acquire these resources efficiently traces back to the evolution of three basic organs: roots, stems, and leaves.
A
root is an organ that anchors a vascular plant in the soil, absorbs mineral and water, and often stores carbohydrates. Most plants have a taproot system, consisting of one main vertical root, the taproot, which develops from the embryonic root and lateral roots, also known as branch roots. Taproot systems generally penetrate deeply and are well adapted to deep soils, where groundwater is not close to the surface. Some roots also form the fibrous root system, a mat of generally thin roots spreading out below the soil surface. Fibrous root systems consist of adventitious roots, roots that arise from unusual location instead of the embryonic roots, and lateral roots branching from each of the small roots. They do not usually penetrate deeply and are best adapted to shallow soils or regions where rainfall is light and does not moisten the soil much below the surface layers. Although the root system helps anchor a plant, absorption of water and minerals in most plants occur primarily near the tips of roots, where vast numbers of root hairs emerge and increase the surface area of the root enormously. Although not an organ, root hairs contribute little to anchorage and its primary function is absorption.
A
stem is an organ that raises or separates leaves, exposing them to sunlight, and reproductive structures, facilitating dispersal of pollen and fruit. Each stem consists of an alternating system of nodes, the points at which leaves are attached, and internodes, the stem segments between nodes. In the upper angle formed by each leaf and stem is an axillary bud, a structure that can form a lateral shoot, or a branch. Most of the growth of a young shoot is concentrated near the shoot tip, which consists of an apical bud, that is composed of developing leaves and a compact series of nodes and internodes. Removal of the apical bud stimulates growth of axillary buds, resulting in more branches.
A
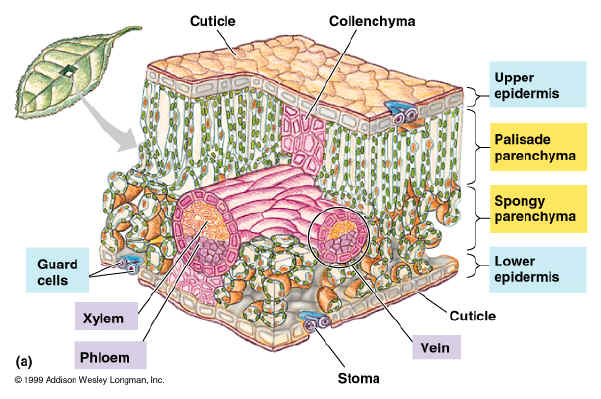
leaf is the main photosynthetic organ in most vascular plants. Leaves vary extensively in form but generally consist of a flattened blade and a stalk, the petioles, which joins the leaf to the stem at a node. Leaves also differ in the arrangement of veins, the vascular tissue of leaves. Some have parallel major veins that run the length of the blade, while others have a branched network of major veins. In identifying plants according to structure, taxonomists rely on variations in leaf morphology, such as leaf shape, the branching pattern of veins, and the spatial arrangement of leaves.
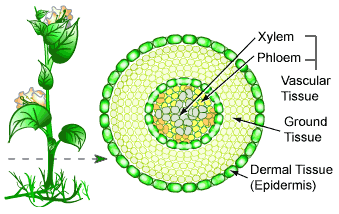
Each plant organ has dermal, vascular and ground tissues, each a part of the plant tissue system and have different functions. The
dermal tissue system is the plant’s outer protective covering. It forms the first line of defense against physical damage and pathogens. In nonwoody plants, it is usually a single tissue called the epidermis, a layer of tightly packed cells. In leaves and most stems, the cuticle, a waxy coating on the epidermal surface, helps prevent water loss. In wood plants, protective tissues called periderm replace the epidermis in older regions of stems and roots. In addition to protecting the plant against water loss and disease, some plants have epidermis that contains trichomes, hairlike outgrowths that provide defense against insects by forming a barrier or by secreting sticky fluids or toxic compounds. The
vascular tissue system carries out long-distance transport of materials between the roots and shoot systems. The two types of vascular tissues are xylem and phloem. Xylem conducts water and dissolved minerals upward from roots into the shoots. Phloem transports sugars, the product of photosynthesis, from where they are produced to where they are needed. The vascular tissue of a root and stem is collectively called the stele and its arrangement varies, depending on the species and organs. Tissues that are neither dermal nor vascular are part of the
ground tissue system. It is responsible for most of the plant’s metabolic functions and is located between the dermal tissue and vascular tissue in each organ.
There are many different types of plant cells:
parenchyma cells, collenchyma cells, sclerenchyma cells, water-conducting cells of the xylem, and
sugar-conducting cells of the phloem. Parenchyma cells have primary walls that are relatively thin and flexible, but lack secondary walls. They perform most of the metabolic functions of the plant, synthesizing and storing various organic products. Most parenchyma cells retain the ability to divide and differentiate into other types of plant cells under particular conditions, such as wound repairs. Grouped in strands, collenchyma cells are generally elongated cells that have thicker, though more uneven, primary walls than parenchyma cells. They provide flexible support without restraining growth and elongate with the stems and leaves they support as they reach maturity. Sclerenchyma cells also function as supporting elements in plants. They are more rigid than collenchyma cells and its secondary wall contain large amount of lignin, indigestible polymer that accounts for more than a quarter of the dry mass of wood. Mature sclerenchyma cells are dead at functional maturity, but their rigid walls remain as “skeletons” that can support the plant for hundreds of years. Two types of sclerenchyma cells are specialized entirely support and strengthening: sclereids and fibers. Sclereids, which are more irregular in shape, have very thick, lignified walls, while fibers, which are usually grouped in strands, are long, slender, and tapered. The xylem has two types of water-conducting cells: tracheids and vessel elements. Both are tubular, elongated cells that are dead at functional maturity and have pits where water can migrate laterally between neighboring cells. The phloem has long, narrow sugar-conducting cells called sieve cells that are alive at functional maturity. The sieve tubes have sieve plates and pores that facilitate the flow of fluid from cell to cell.
Penny is a typical flowering tree plant. She has all three organs necessary for its function. With a functioning taproot system, she can easily absorb minerals and water and store carbohydrates that can will transported throughout the plant. She has a stem to hold her structure up and leaves to conduct photosynthesis. Because she is very sturdy and well-adapt to her environment, Penny must have a tough dermal tissue system and vascular tissue system.








{kind=link}